Il spesso citato "dogma centrale di biologia molecolare" è catturato nel semplice schema Da DNA a RNA a proteine. Leggermente ampliato, questo significa che il acido desossiribonucleico, che è il materiale genetico nel nucleo delle cellule, viene utilizzato per creare una molecola simile chiamata RNA (acido ribonucleico) in un processo chiamato trascrizione. Fatto ciò, l'RNA viene utilizzato per dirigere la sintesi delle proteine altrove nella cellula in un processo chiamato traduzione.
Ogni organismo è la somma delle proteine che produce, e in tutto ciò che è vivo oggi e mai conosciuto hanno vissuto, le informazioni per produrre queste proteine sono immagazzinate in, e solo in, quell'organismo organism DNA. Il tuo DNA è ciò che ti rende ciò che sei ed è ciò che trasmetti a tutti i bambini che potresti avere.
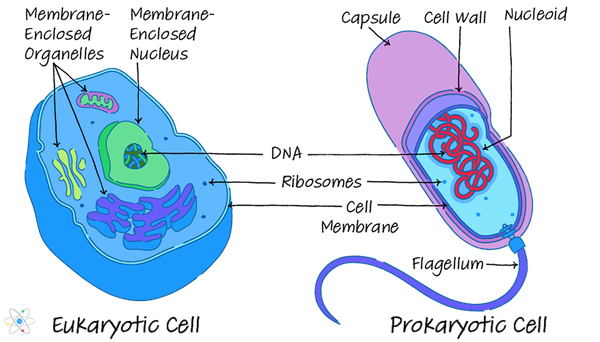
Nel eucariotico organismi, dopo che la prima fase della trascrizione è completa, l'RNA messaggero (mRNA) appena sintetizzato deve trovare la sua strada al di fuori del nucleo nel citoplasma dove avviene la traduzione. (Nei procarioti, che mancano di nuclei, questo non è il caso.) Poiché la membrana plasmatica che circonda il contenuto del nucleo può essere esigente, questo processo richiede un input attivo dalla cellula stessa.
Acidi nucleici
Due acidi nucleici esistono in natura, DNA e RNA. Gli acidi nucleici sono macromolecole in quanto sono composti da catene molto lunghe di subunità ripetute, o monomeri, chiamate nucleotidi. Nucleotidi essi stessi sono costituiti da tre distinti componenti chimici: uno zucchero a cinque atomi di carbonio, da uno a tre gruppi fosfato e una delle quattro basi ricche di azoto (azotate).
Nel DNA, la componente dello zucchero è desossiribosio, mentre nell'RNA è ribosio. Questi zuccheri differiscono solo per il fatto che il ribosio trasporta un gruppo ossidrile (-OH) attaccato a un carbonio al di fuori dell'anello a cinque membri dove il desossiribosio trasporta solo un atomo di idrogeno (-H).
I quattro possibili basi azotate nel DNA sono adenina (A), citosina (C), guanina (G) e timina (T). L'RNA ha i primi tre, ma include uracile (U) al posto della timina. Il DNA è a doppio filamento, con i due filamenti collegati alle loro basi azotate. A si accoppia sempre con T e C si accoppia sempre con G. I gruppi zucchero e fosfato creano la spina dorsale" di ogni cosiddetto filo complementare. La formazione risultante è una doppia elica, la cui forma è stata scoperta negli anni '50.
- Nel DNA e nell'RNA, ogni nucleotide contiene un singolo gruppo fosfato, ma i nucleotidi liberi spesso ne hanno due (ad es. ADP o adenosina difosfato) o tre (ad es. ATP o adenosina trifosfato).
Sintesi dell'RNA messaggero: trascrizione
La trascrizione è la sintesi di una molecola di RNA chiamata RNA messaggero (mRNA), da uno dei filamenti complementari di una molecola di DNA. Esistono anche altri tipi di RNA, il più comune è tRNA (RNA di trasferimento) e RNA ribosomiale (rRNA), entrambi i quali giocano un ruolo critico nella traduzione al ribosoma.
Lo scopo dell'mRNA è creare un insieme di direzioni mobili e codificate per la sintesi delle proteine. Una lunghezza di DNA che include il "progetto" per un singolo prodotto proteico è chiamata gene. Ogni sequenza di tre nucleotidi porta le istruzioni per produrre un particolare amminoacido, con ammino gli acidi sono i mattoni delle proteine nello stesso modo in cui i nucleotidi sono i mattoni dei nuclei acidi.
Ci sono 20 aminoacidi in tutto, consentendo un numero sostanzialmente illimitato di combinazioni e quindi di prodotti proteici.
La trascrizione avviene in nucleo, lungo un singolo filamento di DNA che è stato disaccoppiato dal suo filamento complementare per scopi di trascrizione. Gli enzimi si attaccano alla molecola del DNA all'inizio del gene, in particolare l'RNA polimerasi. L'mRNA che viene sintetizzato è complementare al filamento di DNA utilizzato come stampo, e quindi assomiglia al filamento stampo proprio filamento di DNA complementare tranne che U appare nell'mRNA ovunque T sarebbe apparso se la molecola in crescita DNA anziché.
Trasporto di mRNA all'interno del nucleo Nu
Dopo che le molecole di mRNA sono state sintetizzate nel sito di trascrizione, devono compiere il loro viaggio verso i siti di traduzione, i ribosomi. ribosomi appaiono sia liberi nel citoplasma cellulare che attaccati ad un organello membranoso chiamato reticolo endoplasmatico, entrambi i quali giacciono al di fuori del nucleo.
Prima che l'mRNA possa passare attraverso la doppia membrana plasmatica che costituisce l'involucro nucleare (o membrana nucleare), deve raggiungere in qualche modo la membrana. Ciò si verifica dal legame delle nuove molecole di mRNA per trasportare le proteine.
Prima che i complessi mRNA-proteina (mRNP) risultanti possano spostarsi verso il bordo, si mescolano completamente all'interno della sostanza del nucleo, in modo che quei complessi mRNP che si formano vicino al bordo del nucleo non hanno migliori possibilità di uscire dal nucleo in un dato momento dopo la formazione rispetto ai processi mRNP vicino al nucleo interno.
Quando i complessi mRNP incontrano regioni del nucleo pesanti nel DNA, che in questo ambiente esiste come cromatina (cioè DNA legato a proteine strutturali), può andare in stallo, proprio come un camioncino impantanato in pesanti fango. Questo stallo può essere superato dall'apporto di energia sotto forma di ATP, che spinge l'mRNP impantanato nella direzione del bordo del nucleo.
Complessi di pori nucleari
Il nucleo deve proteggere l'importantissimo materiale genetico della cellula, ma deve anche avere un mezzo per scambiare proteine e acidi nucleici con il citoplasma cellulare. Ciò si ottiene tramite "porte" costituite da proteine e note come complessi di pori nucleari (NPC). Questi complessi hanno un poro che attraversa la doppia membrana dell'involucro nucleare e una serie di strutture diverse su entrambi i lati di questa "porta".
L'NPC è enorme per gli standard molecolari. Negli esseri umani ha una massa molecolare di 125 milioni di Dalton. Al contrario, una molecola di glucosio ha una massa molecolare di 180 Dalton, che la rende circa 700.000 volte più piccola del complesso NPC. Sia il trasporto dell'acido nucleico che quello delle proteine nel nucleo e il movimento di queste molecole fuori dal nucleo avvengono tramite l'NPC.
Sul lato citoplasmatico, l'NPC ha quello che viene chiamato un anello citoplasmatico e filamenti citoplasmatici, entrambi i quali servono per aiutare ad ancorare l'NPC in posizione nella membrana nucleare. Sul lato nucleare dell'NPC c'è un anello nucleare, analogo all'anello citoplasmatico sul lato opposto, così come un cesto nucleare.
Una varietà di singole proteine partecipano al movimento dell'mRNA e una varietà diversificata di altre proteine carichi molecolari fuori dal nucleo, con lo stesso che si applica al movimento di sostanze nel nucleo.
Funzione mRNA nella traduzione
L'mRNA non inizia il suo lavoro effettivo finché non raggiunge un ribosoma. Ogni ribosoma nel citoplasma o attaccato a reticolo endoplasmatico consiste in una subunità grande e una piccola; questi si uniscono solo quando il ribosoma è attivo nella trascrizione.
Quando una molecola di mRNA si attacca ad a traduzione sito lungo il ribosoma, è unito da un particolare tipo di tRNA che trasporta uno specifico amminoacido (esistono quindi 20 gusti diversi di tRNA, uno per ogni amminoacido). Ciò si verifica perché il tRNA può "leggere" la sequenza di tre nucleotidi sull'mRNA esposto che corrisponde a un dato amminoacido.
Quando il tRNA e l'mRNA "si accoppiano", il tRNA rilascia il suo amminoacido, che viene aggiunto all'estremità della catena di amminoacidi in crescita destinata a diventare una proteina. Questo polipeptide raggiunge la lunghezza specificata quando la molecola di mRNA viene letta nella sua interezza e il polipeptide viene rilasciato e trasformato in una proteina autentica.