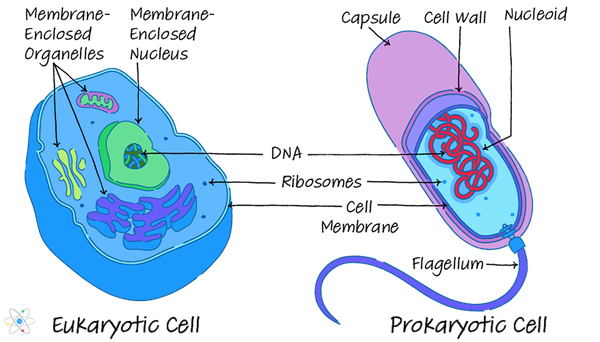
Respirația aerobă, un termen folosit adesea în mod interschimbabil cu „respirația celulară”, este un mod minunat de randament extrageți energia stocată în legăturile chimice ale compușilor de carbon în prezența oxigenului și folosiți această energie extrasă pentru a o folosi în metabolică proceselor. Organismele eucariote (adică animale, plante și ciuperci) folosesc toate respirația aerobă, datorită în principal prezenței organelor celulare numite mitocondrii. Câteva organisme procariote (adică bacterii) folosesc căi de răsuflare aerobică mai rudimentare, dar, în general, când vedeți „respirația aerobă”, ar trebui să credeți „eucariotă multicelulară organism."
Dar nu numai asta ar trebui să vă sară în minte. Ceea ce urmează vă spune tot ce trebuie să știți despre căile chimice de bază ale respirației aerobe, de ce este un astfel de set esențial de reacții și cum a început totul pe parcursul biologic și geologic istorie.
Rezumatul chimic al respirației aerobe
Tot metabolismul celular al nutrienților începe cu molecule de glucoză. Acest zahăr cu șase carbon poate fi derivat din alimente din toate cele trei clase de macronutrienți (carbohidrați, proteine și grăsimi), deși glucoza însăși este un carbohidrat simplu. În prezența oxigenului, glucoza este transformată și descompusă într-un lanț de aproximativ 20 de reacții pentru a produce dioxid de carbon, apă, căldură, și 36 sau 38 de molecule de adenozin trifosfat (ATP), molecula cel mai des utilizată de celule în toate viețuitoarele ca sursă directă de combustibil. Variația cantității de ATP produsă de respirația aerobă reflectă faptul că plantele celulare uneori stoarce 38 de ATP dintr-o moleculă de glucoză, în timp ce celulele animale generează 36 de ATP pe glucoză moleculă. Acest ATP provine din combinarea moleculelor de fosfat liber (P) și adenozin difosfatului (ADP), cu aproape toate acest lucru având loc chiar în ultimele etape ale respirației aerobe în reacțiile transportului de electroni lanţ.
Reacția chimică completă care descrie respirația aerobă este:
C6H12O6 + 36 (sau 38) ADP + 36 (sau 38) P + 6O2 → 6CO2 + 6H2O + 420 kcal + 36 (sau 38) ATP.
În timp ce reacția în sine pare suficient de simplă în această formă, contrazice multitudinea de pași pe care îi ia pentru a obține de la partea stângă a ecuației (reactanții) la partea dreaptă (produsele, inclusiv 420 kilocalorii de căldură). Prin convenție, întreaga colecție de reacții este împărțită în trei părți bazate pe locul în care apar fiecare: glicoliza (citoplasma), ciclul Krebs (matricea mitocondrială) și lanțul de transport al electronilor (mitocondria interioară) membrană). Cu toate acestea, înainte de a explora în detaliu aceste procese, este în ordine o privire asupra modului în care respirația aerobă a început pe Pământ.
Originile sau respirația aerobă a Pământului
Funcția respirației aerobe este de a furniza combustibil pentru repararea, creșterea și întreținerea celulelor și țesuturilor. Acesta este un mod oarecum formal de a observa că respirația aerobă menține în viață organismele eucariote. Ați putea rămâne multe zile fără mâncare și cel puțin câteva fără apă în majoritatea cazurilor, dar doar câteva minute fără oxigen.
Oxigenul (O) se găsește în aerul normal sub forma sa diatomică, O2. Acest element a fost descoperit, într-un anumit sens, în anii 1600, când a devenit evident pentru oamenii de știință că aerul conținea un element vitală pentru supraviețuirea animalelor, una care ar putea fi epuizată într-un mediu închis prin flacără sau, pe termen lung, prin respiraţie.
Oxigenul reprezintă aproximativ o cincime din amestecul de gaze în care respirați. Dar nu a fost întotdeauna așa în istoria planetei de 4,5 miliarde de ani și în schimbarea cantitatea de oxigen din atmosfera Pământului a avut de-a lungul timpului efecte profunde asupra biologice evoluţie. În prima jumătate a vieții actuale a planetei, a existat Nu oxigen din aer. În urmă cu 1,7 miliarde de ani, atmosfera era formată din 4% oxigen și apăruseră organisme unicelulare. Cu 0,7 miliarde de ani în urmă, O2 compus între 10 și 20 la sută din aer și au apărut organisme multicelulare mai mari. În urmă cu 300 de milioane de ani, conținutul de oxigen a crescut la 35% din aer și, în consecință, dinozaurii și alte animale foarte mari erau norma. Mai târziu, cota de aer deținută de O2 a scăzut la 15% până a ridicat din nou locul unde se află astăzi.
Este clar, urmărind doar acest model, care pare extrem de științific că funcția finală a oxigenului este de a face animalele să crească mari.
Glicoliza: un punct de plecare universal
Cele 10 reacții ale glicolizei nu necesită ele însele oxigen pentru a continua și glicoliza apare într-o oarecare măsură în toate viețuitoarele, atât procariote, cât și eucariote. Dar glicoliza este un precursor necesar pentru reacțiile aerobe specifice ale respirației celulare și este descrisă în mod normal împreună cu acestea.
Odată ce glucoza, o moleculă cu șase carbon cu o structură inelară hexagonală, intră în citoplasma unei celule, este fosforilată imediat, ceea ce înseamnă că are o grupare fosfat atașată la unul dintre carbonii săi. Acest lucru captează în mod eficient molecula de glucoză din interiorul celulei, oferindu-i o sarcină netă negativă. Molecula este apoi rearanjată în fructoză fosforilată, fără pierderi sau câștig de atomi, înainte ca încă un fosfat să fie adăugat la moleculă. Acest lucru destabilizează molecula, care apoi se fragmentează într-o pereche de compuși cu trei atomi de carbon, fiecare dintre ei având propriul său fosfat atașat. Una dintre acestea se transformă în cealaltă și apoi, într-o serie de etape, cele două molecule de trei carbon renunță la fosfații lor la molecule de ADP (adenozin difosfat) pentru a produce 2 ATP. Molecula originală de glucoză cu șase carbonuri se înfășoară ca două molecule ale unei molecule cu trei cărbuni numită piruvat și, în plus, sunt generate două molecule de NADH (discutate în detaliu mai târziu).
Ciclul Krebs
Piruvatul, în prezența oxigenului, se deplasează în matricea (gândiți-vă la „mijloc”) a organelor celulare numită mitocondrie și este transformată într-un compus cu doi atomi de carbon, numit acetil coenzima A (acetil CoA). În acest proces, o moleculă de dioxid de carbon (CO2). În acest proces, o moleculă de NAD+ (așa-numitul purtător de electroni de mare energie) este convertit în NADH.
Ciclul Krebs, numit și ciclul acidului citric sau ciclul acidului tricarboxilic, este denumit mai degrabă un ciclu decât o reacție deoarece unul dintre produsele sale, oxaloacetatul de moleculă cu patru carbon, reintră la începutul ciclului prin combinarea cu o moleculă de acetil CoA. Acest lucru are ca rezultat o moleculă de șase carbon numită citrat. Această moleculă este manipulată de o serie de enzime într-un compus cu cinci atomi de carbon numit alfa-cetoglutarat, care apoi pierde un alt carbon pentru a produce succinat. De fiecare dată când se pierde un carbon, acesta este sub formă de CO2, și deoarece aceste reacții sunt favorabile din punct de vedere energetic, fiecare pierdere de dioxid de carbon este însoțită de conversia unui alt NAD+ la NAD. Formarea succinatului creează, de asemenea, o moleculă de ATP.
Succinatul este transformat în fumarat, generând o moleculă de FADH2 din FAD2+ (un purtător de electroni similar cu NAD+ în funcțiune). Acesta este transformat în malat, rezultând un alt NADH, care este apoi transformat în oxaloacetat.
Dacă țineți scorul, puteți număra 3 NADH, 1 FADH2 și 1 ATP pe rândul ciclului Krebs. Dar rețineți că fiecare moleculă de glucoză furnizează două molecule de acetil CoA pentru intrarea în ciclu, astfel încât numărul total al acestor molecule sintetizate este de 6 NADH, 2 FADH2 și 2 ATP. Prin urmare, ciclul Krebs nu generează multă energie în mod direct - doar 2 ATP pe moleculă de glucoză furnizată în amonte - și nu este necesar nici oxigen. Dar NADH și FADH2 sunt critice pentru fosforilarea oxidativă pași în următoarea serie de reacții, denumită în mod colectiv lanțul de transport al electronilor.
Lanțul de transport al electronilor
Diferitele molecule de NADH și FADH2 create în etapele precedente ale respirației celulare sunt gata de utilizare în lanțul de transport al electronilor, care are loc în pliurile membranei mitocondriale interioare numite criste. Pe scurt, electronii de mare energie atașați la NAD+ și FAD2+ sunt folosite pentru a crea un gradient de protoni peste membrană. Aceasta înseamnă doar că există o concentrație mai mare de protoni (H+ ioni) pe o parte a membranei decât pe cealaltă parte, creând un impuls pentru ca acești ioni să curgă din zone cu concentrație mai mare de protoni în zone cu concentrație mai mică de protoni. În acest fel, protonii se comportă puțin diferit față de, să zicem, apa care „vrea” să se deplaseze dintr-o zonă de altitudine mai înaltă într-o zonă de mai mică concentrație - aici, sub influența gravitației în locul așa-numitului gradient chemiosmotic observat în transportul electronilor lanţ.
La fel ca o turbină la o centrală hidroelectrică care valorifică energia apei curgătoare pentru a lucra în altă parte (în acest caz, generează electricitate), o parte din energia stabilită de proton gradientul de-a lungul membranei este capturat pentru a atașa grupări fosfat libere (P) la moleculele ADP pentru a genera ATP, un proces numit fosforilare (și în acest caz, oxidativ fosforilare). De fapt, acest lucru se întâmplă mereu în lanțul de transport al electronilor, până când toate NADH și FADH2 din glicoliză și ciclul Krebs - aproximativ 10 dintre primele și două din cele din urmă - sunt utilizate. Acest lucru are ca rezultat crearea a aproximativ 34 de molecule de ATP pe moleculă de glucoză. Deoarece glicoliza și ciclul Krebs produc fiecare câte 2 ATP pe moleculă de glucoză, cantitatea totală dacă energia eliberată, cel puțin în condiții ideale, este de 34 + 2 + 2 = 38 ATP în total.
Există trei puncte diferite în lanțul de transport al electronilor în care protonii pot traversa membrana mitocondrială interioară pentru a intra în spațiul dintre aceasta ulterior și membrana mitocondrială externă și patru complexe moleculare distincte (numerotate I, II, III și IV) care formează punctele fizice de ancorare ale lanţ.
Lanțul de transport al electronilor necesită oxigen deoarece O2 servește ca acceptor final al perechii de electroni din lanț. Dacă nu este prezent oxigen, reacțiile din lanț încetează rapid, deoarece fluxul „în aval” de electroni încetează; nu au încotro. Printre substanțele care pot paraliza lanțul de transport al electronilor se numără cianura (CN-). Acesta este motivul pentru care este posibil să fi văzut cianura folosită ca otravă mortală în spectacole de omucidere sau filme de spionaj; atunci când este administrat în doze suficiente, respirația aerobă în interiorul destinatarului se oprește și, odată cu aceasta, viața însăși.
Fotosinteza și respirația aerobă la plante
Se presupune adesea că plantele suferă fotosinteza pentru a crea oxigen din dioxid de carbon, în timp ce animalele folosesc respirația pentru a genera dioxid de carbon din oxigen, ajutând astfel la conservarea unui ecosistem îngrijit, complementar echilibru. Deși acest lucru este adevărat la suprafață, este înșelător, deoarece plantele folosesc atât fotosinteza, cât și respirația aerobă.
Deoarece plantele nu pot mânca, ele trebuie să facă, mai degrabă decât să ingereze, hrana lor. Pentru aceasta este destinată fotosinteza, o serie de reacții care au loc la organele cărora le lipsește animalele numite cloroplaste. Alimentat de lumina soarelui, CO2 în interiorul celulei vegetale este asamblat în glucoză în interiorul cloroplastelor într-o serie de pași care seamănă cu lanțul de transport al electronilor din mitocondrii. Glucoza este apoi eliberată din cloroplast; cel mai mult dacă devine o porțiune structurală a plantei, dar unele suferă glicoliză și apoi continuă prin restul respirației aerobe după intrarea în mitocondriile celulei plantei.